








{kind=link}
{kind=link}
{kind=link}
{kind=link}
INTRODUCTION
Plasmodium falciparum and Plasmodium vivax are the predominant parasites responsible for human malaria worldwide, contributing nearly 241 million clinical cases in 2020.1 Given their contribution to the global burden of malaria, P. falciparum and P. vivax remain the primary focus of public health efforts aimed at achieving malaria control, elimination, and eradication.2 However, three additional major Plasmodium species—P. knowlesi, P. malariae, and P. ovale—also cause clinical episodes of malaria, although their overall burden remains uncertain due in part to routine misdiagnosis in a clinical setting.3
Diagnosis of clinical malaria is routinely performed using light microscopy (LM). In addition to challenges presented by low or submicroscopic parasite densities,4 accurate speciation of malaria parasites using LM depends on identification of morphological features characteristic of the Plasmodium spp. causing the infection. However, morphological similarities across the five Plasmodium spp., such as the resemblance of early blood-stage P. knowlesi parasites to P. falciparum parasites and late blood-stage P. knowlesi parasites to P. malariae parasites,2,5 undermine species-level identification of Plasmodium infections.6
In settings with co-circulation of the Plasmodium spp., routine misdiagnosis by light microscopy has consequences for clinical decision-making and malaria control. The choice of therapeutic is specific to the Plasmodium spp. causing the infection,7 so misdiagnosis may lead to ineffective treatment of blood-stage parasites for potentially life-threatening Plasmodium spp. infections as well as ineffective or no treatment of liver-stage parasites for P. vivax and P. ovale infections. Additionally, misdiagnosis may present challenges for monitoring Plasmodium spp. transmission, particularly for the emerging zoonosis P. knowlesi. Currently, the extent of human-to-human transmission of P. knowlesi is unknown,2,8 and misdiagnosis could cause transmission clusters to appear smaller than their actual size, leading to an underestimate of transmission.9
Consequently, measuring the extent of misdiagnosis by light microscopy is of importance to public health, and past studies have directly compared the diagnostic performance of light microscopy to polymerase chain reaction (PCR) in settings with co-circulation of Plasmodium spp. parasites.6,10–21 However, these studies varied in their criteria for enrollment and other study design features, as well as in the epidemiological settings in which they were conducted. This makes direct comparison of their estimates of diagnostic performance challenging.
To synthesize across these studies, we performed a systematic review and meta-analysis of the diagnostic performance of light microscopy in settings with co-circulation of all five Plasmodium spp. We identified studies that compared the diagnostic performance of light microscopy to PCR and fit a hierarchical Bayesian latent class model to estimate study- and species-level sensitivities and specificities of light microscopy. Finally, to assess whether the perceived prevalences of the five species might affect diagnostic outcomes of light microscopy, we explored the relationship between the estimates of sensitivity and specificity that we obtained and the observed PCR prevalences in each study.
METHODS
Search strategy and selection criteria.
In this systematic review and meta-analysis, JHH searched MEDLINE and Web of Science for studies published in English before April 30, 2021, that compared the diagnostic performance of LM and PCR for Plasmodium spp. infections. We identified studies using the following search strategy: (Plasmodium OR malaria) AND knowlesi AND ((microscopy OR microscopic OR blood film OR thick film OR thin film) AND (PCR or “polymerase chain reaction”)).
Studies met the criteria for inclusion if they compared the performance of LM and PCR on clinical samples collected in areas with endemic transmission of all five Plasmodium spp. that naturally infect humans (i.e., P. falciparum, P. vivax, P. knowlesi, P. malariae, and P. ovale). Furthermore, studies were only included if they used a PCR protocol that was capable of amplifying DNA from all five Plasmodium spp. Before 2004, the public health burden of P. knowlesi was underappreciated,5 so restricting our search to studies published between 2004 and 2021 ensured that the diagnostic outcomes were representative of the current clinical and diagnostic landscape.
The data were extracted from each study that met the criteria for inclusion. Because the data in each study were anonymized and reported in aggregate, ethical approvals were not required according to a determination from the University of Notre Dame Institutional Review Board that this study did not constitute human subjects research.
Data analysis.
Overview.
In Eq. (1),
A key assumption of the latent class model is that diagnostic outcomes are conditionally independent. That is, by assuming that the sensitivity and specificity of a given diagnostic method is unique for each Plasmodium spp., the outcome that we observe for one parasite species using a given diagnostic method is modeled as independent of the outcomes observed for all other parasite species and diagnostic methods. This implies that the probability of a set of diagnostic outcomes observed across all five Plasmodium spp. is equal to the product of the probabilities of the diagnostic outcome for each Plasmodium spp. Additionally, the probability of acquiring a mixed infection is equal to the product of the prevalence of each Plasmodium spp. present within that mixed infection. Nevertheless, the assumption of conditional independence still allows for interactions between Plasmodium spp. that arise in the context of misdiagnosis. For example, if a P. knowlesi infection is misdiagnosed as a P. malariae infection using LM, then the sensitivity of LM for P. knowlesi and the specificity of LM for P. malariae are penalized because both a false-negative P. knowlesi diagnosis and a false-positive P. malariae diagnosis occurred.
Accounting for differences in study design.
Studies identified in the systematic review varied by enrollment and diagnostic criteria. Where enrollment in a study required a positive Plasmodium spp. diagnosis by LM or PCR, we normalized the probabilities of Eq. (1), such that they were consistent with the subset of possible diagnostic outcomes that could be observed with that study design.
Because P. knowlesi and P. malariae share morphological similarities,2,5 studies may diagnose samples as “P. knowlesi/P. malariae” with LM, indicating that the microscopist identified P. knowlesi and/or P. malariae parasites in the sample but could not make a more precise monoinfection or coinfection diagnosis. To accommodate this in our model, we defined parameters
Inference.
Using PCR as the gold standard, we fit a hierarchical latent class model of clinical diagnosis to the studies identified in our systematic review to estimate study-level and species-level variation in LM sensitivity and specificity for each Plasmodium spp. We fit our model in a Bayesian framework using Stan by running four independent chains of 2,000 samples, each with a warm-up period of 1,000 samples.23 We tested for convergence with the Gelman–Rubin statistic and pooled the independent chains for a posterior distribution of 4,000 samples. Supplementary analyses were performed to evaluate the sensitivity of our posterior estimates to the inclusion of each study and to the assumption of PCR as the gold standard. See the supplemental information for more details.
RESULTS
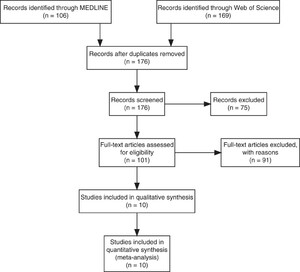
Of the 176 unique studies identified using MEDLINE and Web of Science, 10 studies met the inclusion criteria, and their data comparing LM and PCR diagnostic performance were extracted (Figure 1). The studies enrolled febrile patients presenting in health clinics in Malaysia (N = 6), Thailand (N = 2), Indonesia (N = 1), Myanmar (N = 1), and China (N = 1). Among the 7,999 total patients enrolled across the 12 studies, the pooled percentage positive by PCR was 19% for P. falciparum, 25% for P. vivax, 52% for P. knowlesi, 1.1% for P. malariae, and 0.48% for P. ovale (Table 1). The studies varied widely in the percentage positive by PCR across Plasmodium spp., encompassing a wide range of epidemiological settings and study designs (i.e., criteria for patient enrollment) in which we estimated LM diagnostic performance.

Preferred Reporting Items for Systematic Reviews flow diagram. Flow diagram of the studies identified using MEDLINE and Web of Science for the systematic review.
Citation: The American Journal of Tropical Medicine and Hygiene 108, 1; 10.4269/ajtmh.21-1155
Reported Plasmodium spp. percentage positive by diagnostic method. The percentage of study participants positive by light microscopy (LM) and polymerase chain reaction (PCR) are reported for each Plasmodium spp. in each study, along with the study location, year, and total number and type of samples collected. For P. knowlesi and P. malariae LM percentage of study participants positive.
| Ref. | Location | Samples | Type | Years | P. falciparum | P. vivax | P. knowlesi | P. malariae | P. ovale | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCR | LM | PCR | LM | PCR | LM | PCR | LM | PCR | LM | |||||
| 18 | Thailand | 1,874 | Any Febrile | 2006–2007 | 41 (762) | 37 (687) | 64 (1192) | 54 (1021) | 0.53 (10) | 0.0 (0) | 1.3 (24) | 0.16 (3) | 0.96 (18) | 0.0 (0) |
| 15 | Malaysia | 189 | LM+ | 2008–2011 | 37 (70) | 39 (74) | 35 (66) | 35 (67) | 22 (42) | 0.0 (0) | 1.1 (2) | 27 (51) | 0.0 (0) | 0.0 (0) |
| 10 | Malaysia | 461 | LM+ | 2009–2011 | 7.2 (33) | 12 (54) | 16 (74) | 13 (59) | 85 (392) | 92* (425) | 0.87 (4) | 2* (425) | 0.0 (0) | 0.0 (0) |
| 12 | Malaysia | 227 | Any Febrile | 2008–2010 | 42 (95) | 41 (92) | 41 (92) | 38 (87) | 16 (36) | 0.0 (0) | 0.88 (2) | 15 (33) | 0.044 (1) | 0.044 (1) |
| 20 | China | 560 | Any Febrile | 2008–2012 | 28 (159) | 28 (156) | 51 (288) | 49 (276) | 0.36 (2) | 0.0 (0) | 1.6 (9) | 0.0 (0) | 2.7 (15) | 0.0 (0) |
| 6 | Malaysia | 303 | PCR+/LM+ | 2010–2011 | 43 (130) | 47 (142) | 16 (47) | 16 (47) | 44 (132) | 42* (128) | 0.66 (2) | 42* (128) | 0.0 (0) | 0.0 (0) |
| 11 | Thailand | 297 | Any Febrile | 2012 | 9.1 (27) | 3.4 (10) | 8.8 (26) | 5.1 (15) | 0.0 (0) | 0.0 (0) | 0.34 (1) | 0.34 (1) | 0.0 (0) | 0.0 (0) |
| 19 | Malaysia | 457 | LM+ | 2012–2013 | 11 (51) | 12 (54) | 32 (144) | 30 (138) | 58 (267) | 40 (185) | 0.22 (1) | 18 (81) | 0.044 (2) | 0.022 (1) |
| 16 | Myanmar | 90 | Any Febrile | 2013-2015 | 44 (40) | 43 (39) | 20 (18) | 19 (17) | 0.0 (0) | 0.0 (0) | 1.1 (1) | 1.1 (1) | 0.0 (0) | 0.0 (0) |
| 13 | Malaysia | 3,541 | PCR+/LM+ | 2015–2017 | 4.9 (173) | 4.9 (174) | 2.5 (87) | 2.5 (87) | 92 (3255) | 93* (3280) | 1.1 (40) | 93* (3280) | 0.085 (3) | 0.0 (0) |
| Total | 7,999 | 19 (1,540) | 19 (1,482) | 25 (2,034) | 23 (1,814) | 52 (4,136) | 50 (4,018) | 1.1 (86) | 50 (4,003) | 0.48 (39) | 0.025 (2) | |||
The percentage positive was computed using a diagnosis of P. knowlesi and P. malariae.
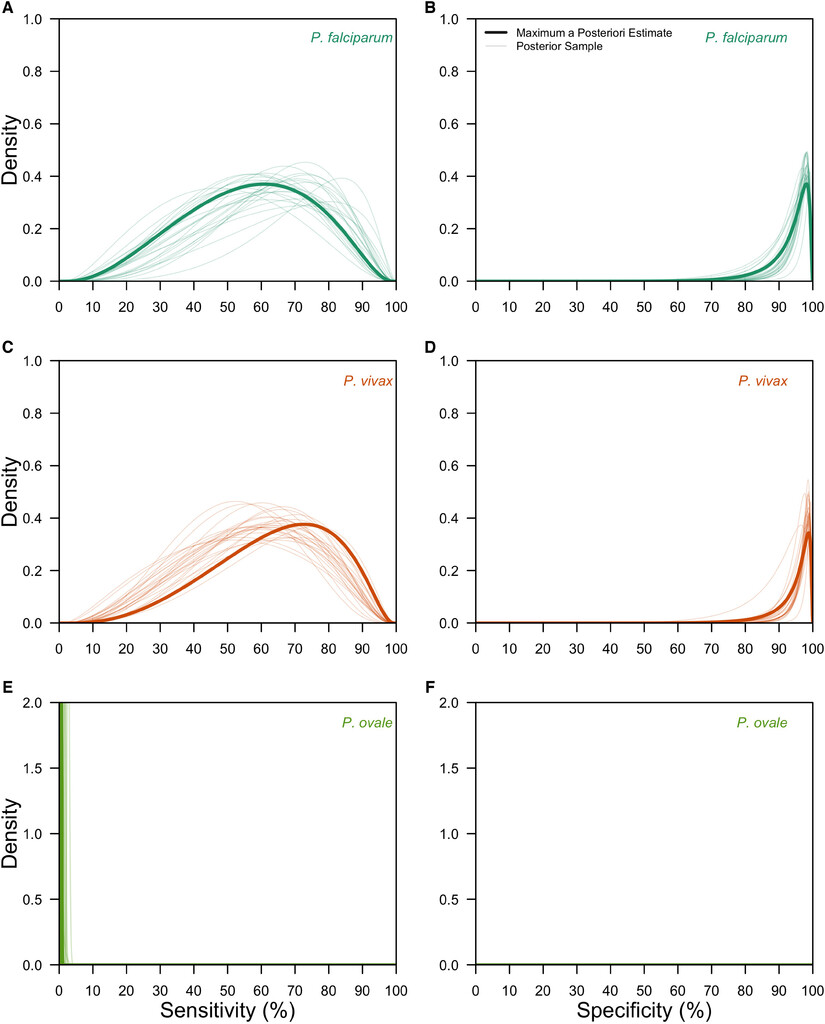
After validating on 200 simulated datasets (Supplemental Figure 1), we applied our hierarchical latent class model to the data from the 10 studies identified in the systematic review. Our fitted hierarchical latent class model captured the variation in the observed data (Supplemental Figure 6) and revealed differences in LM diagnostic performance across Plasmodium spp. (Figure 2). Specifically, we estimated that the species-level mean sensitivity of LM was 65.7% (95% confidence interval [CI]: 48.1–80.3%) for P. falciparum, 62.7% (95% CI: 46.0–77.2%) for P. vivax, and 0.525% (95% CI: 0.0210–3.11%) for P. ovale. It was not possible to estimate group-level mean sensitivities for the pooled P. knowlesi and P. malariae LM diagnosis due to identifiability concerns, yet the mean of study-level posterior median estimates were 44.9% for P. knowlesi parasites and 45.3% for P. malariae parasites. Species-level mean specificity of LM was consistently high across Plasmodium spp. and was estimated at 98.2% (95% CI: 96.6–99.1%) for P. falciparum, 98.6% (95% CI: 97.3–99.3%) for P. vivax, and 100% (95% CI: 100%–100%) for P. ovale. For the pooled P. knowlesi and P. malariae LM diagnosis, the mean of study-level posterior median estimates of specificity were 95.6% for P. knowlesi parasites and 95.9% for P. malariae parasites. These results were robust to the assumption of PCR as the gold standard (Supplemental Figure 9).
Group-level posterior estimates of light microscopy (LM) diagnostic performance. The group-level distributions of LM sensitivity (A, C, E) and LM specificity (B, D, F) are shown for Plasmodium falciparum (teal; A, B), Plasmodium vivax (orange; C, D), and Plasmodium ovale (green; E, F). Thick lines are the maximum a posteriori estimates, and thin lines are 25 posterior samples.
Citation: The American Journal of Tropical Medicine and Hygiene 108, 1; 10.4269/ajtmh.21-1155
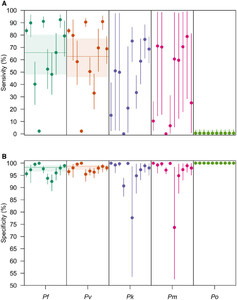
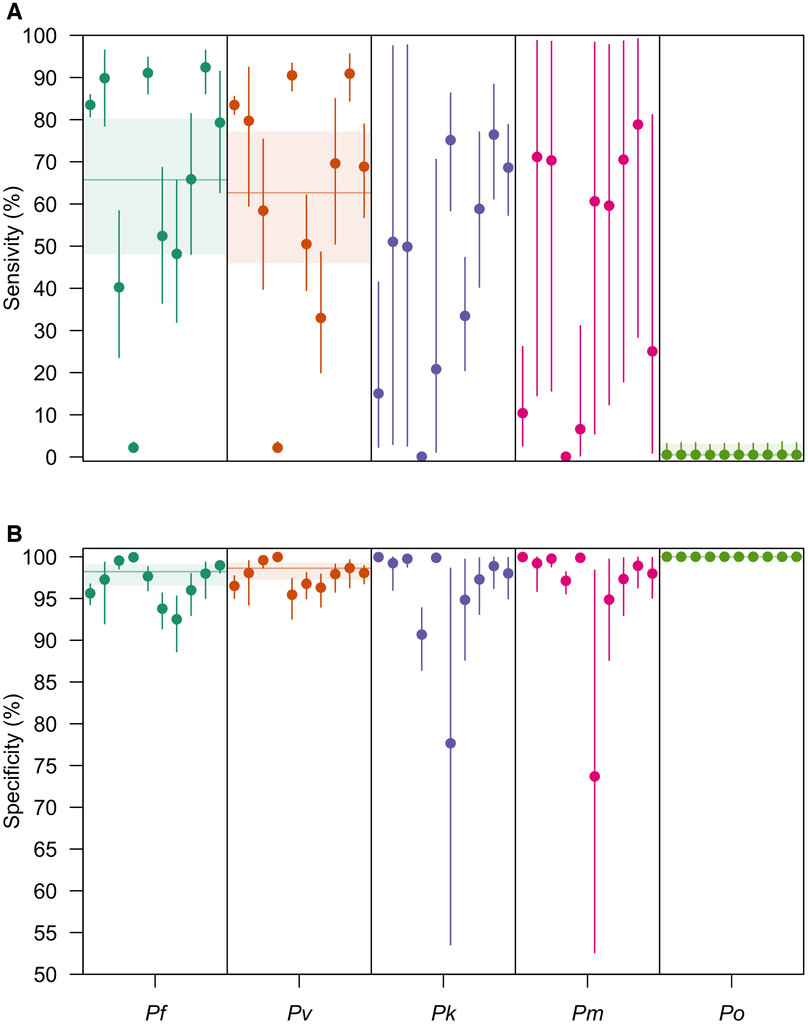
The variance in the hierarchical distributions of LM diagnostic performance in Figure 2 revealed the extent of variation across studies, a feature that depended on both the diagnostic measure and the Plasmodium spp. considered (Figure 3). In general, for a given Plasmodium spp., we estimated greater variation across studies in LM sensitivity than specificity. For example, for P. falciparum, we estimated that logit-transformed standard deviation of LM sensitivity was 1.09 (0.886–1.36), suggesting that the range of sensitivities that fell within one standard deviation of the mean was 60.9% to 85.1%. By comparison, the estimated logit-transformed standard deviation of LM specificity for P. falciparum was 0.967 (0.753–1.23), corresponding to a range of specificities within one standard deviation of the mean of 95.4% to 99.3%. Across Plasmodium spp., study-level estimates of LM sensitivity and specificity for P. falciparum, P. vivax, P. knowlesi, and P. malariae exhibited greater variation than for P. ovale.
Site-level posterior estimates of light microscopy (LM) diagnostic performance. The site-level posterior estimates of (A) LM sensitivity and (B) LM specificity are shown for P. falciparum (teal), Plasmodium vivax (orange), Plasmodium knowlesi (purple), Plasmodium malariae (pink), and Plasmodium ovale (green). Circles are the median posterior estimate, and the vertical segment is the 95% credible interval. The horizontal line is the posterior median of the group-level mean, and the horizontal shaded region is the corresponding 95% credible interval.
Citation: The American Journal of Tropical Medicine and Hygiene 108, 1; 10.4269/ajtmh.21-1155
Because the estimates of LM diagnostic performance varied across the unique studies, we then assessed its relationship with observed PCR prevalence (Figure 4). We observed strong, positive correlations between PCR prevalence and LM sensitivity for P. vivax (Pearson correlation test: ρ = 0.78; P = 0.0082) and P. falciparum (ρ = 0.62; P = 0.054) and weaker, positive correlations for P. knowlesi (ρ = 0.01; P = 0.97) and P. ovale (ρ = 0.06; P = 0.087), although the latter two were not statistically significant associations. By contrast, for P. malariae, there was a negative correlation between PCR prevalence and LM sensitivity, although this estimate was also not statistically significant. Across Plasmodium spp., increased PCR prevalence of P. knowlesi was associated with reduced sensitivity of LM for P. falciparum (ρ = –0.65; P = 0.042) and P. vivax (ρ = –0.76; P = 0.010), highlighting a possible source of misdiagnosis. Additional negative associations were observed between PCR prevalence and LM sensitivity across P. falciparum, P. knowlesi, and P. malariae—Plasmodium spp. that are commonly misdiagnosed, although these quantities were not statistically significant.
Correlations between Plasmodium spp. fever prevalence and light microscopy (LM) diagnostic performance. The correlations between Plasmodium spp. fever prevalence and (A) LM sensitivity and (B) LM specificity are reported. The color represents the strength of the correlation, with red colors denoting positive correlations and blue colors denoting negative correlations. * P < 0.05, ** P < 0.01, and *** P < 0.001.
Citation: The American Journal of Tropical Medicine and Hygiene 108, 1; 10.4269/ajtmh.21-1155
Generally, for a given Plasmodium spp., PCR prevalence was negatively associated with LM specificity (Figure 4B). For P. knowlesi, this association was strong and statistically significant (ρ = –0,79; P = 0.0061), and additional weaker, yet statistically nonsignificant relationships were observed for P. falciparum, P. vivax, and P. ovale. The opposite relationship was observed for P. malariae, although this quantity was not statistically significant. Across Plasmodium spp., increased prevalence of P. malariae was associated with reduced specificities of LM for P. falciparum and P. vivax, and increased prevalence of P. knowlesi was associated with reduced specificity of LM for P. malariae, though these effects were not statistically significant.
DISCUSSION
In this systematic review and meta-analysis, we estimated study- and species-level variation in the malaria diagnostic performance of light microscopy in epidemiological settings with cocirculation of the five major Plasmodium spp. that cause human malaria. Our analysis revealed variable sensitivity of light microscopy to diagnose Plasmodium spp. infection. Sensitivity of light microscopy was estimated to be considerably lower for P. knowlesi, P. malariae, and P. ovale than for P. falciparum and P. vivax. This suggests that the burden of P. knowlesi, P. malariae, and P. ovale in a clinical setting may be appreciably underestimated, consistent with a recent study of P. ovale in health clinics in Kenya.3
Post hoc analyses revealed relationships between the diagnostic performance of light microscopy and the epidemiological setting. In general, we noted a positive association between the PCR prevalence of a Plasmodium spp. and the sensitivity of light microscopy for that Plasmodium spp. This supports a phenomenon whereby the more a microscopist encounters samples of a Plasmodium spp. infection in a clinical setting, the more likely they are to accurately identify the Plasmodium spp. causing that infection. By contrast, increased PCR prevalence of a Plasmodium spp. was associated with high false-positive probabilities for that Plasmodium spp. This tendency to misdiagnose malaria infections as what is most commonly encountered in that clinical setting could lead to ineffective monitoring of less commonly encountered Plasmodium spp., such as P. knowlesi, P. malariae, and P. ovale, and especially could present challenges in settings where a Plasmodium spp. is newly emerging.
The high probabilities of misdiagnosis estimated in this analysis could have important consequences for malaria treatment and control. Specifically, we observed an association between increased prevalence of P. knowlesi and reduced sensitivity of LM for P. vivax. This suggests that in areas with P. knowlesi, infections of P. vivax may not be correctly diagnosed and therefore may not receive radical cure therapy to clear hypnozoites and prevent relapse.24 Additionally, increased prevalence of P. malariae was associated with reduced specificity of LM for P. vivax, indicating that many P. malariae infections may be misdiagnosed as P. vivax. This form of misdiagnosis could unnecessarily subject individuals to radical cure therapy, potentially increasing the risk of acute hemolysis among individuals deficient of the glucose-6-phosphate dehydrogenase enzyme.25 The potential for misdiagnosis in the setting of cocirculation should thus be a consideration when guiding treatment recommendations, although this will likely depend strongly on local epidemiology.
Routine diagnosis of clinical malaria by LM may prevent effective monitoring of P. knowlesi transmission and burden. We estimated variable sensitivity of LM to diagnose P. knowlesi and P. malariae infections, given the morphological similarities of P. falciparum, P. knowlesi, and P. malariae parasites2,5 as well as likely delays in familiarity of P. knowlesi among microscopists in the health systems. Although in practice P. knowlesi and P. malariae infections diagnosed by light microscopy are confirmed by PCR in many settings,10 P. knowlesi infections misdiagnosed as P. falciparum, for instance, would not be subject to confirmatory PCR. This could lead to an underestimate of P. knowlesi burden, affecting epidemiological assessments of P. knowlesi infection risk.
Our analysis is subject to several limitations. Our systematic review may not have identified all studies with relevant data, including those studies not indexed by MEDLINE and Web of Science. We limited our review to studies that tested for all five Plasmodium spp. However, studies that included subsets of the five Plasmodium spp. could still be informative of species-level diagnostic performance. Our latent class model assumed conditional independence of diagnosis across Plasmodium spp. Future extensions of this work could account for the correlation structure of LM diagnosis across the Plasmodium spp., due to shared morphologies of the parasites. Additionally, due to the data available for this analysis, it was not possible to estimate sensitivity and specificity as a function of Plasmodium parasitemia. Future work could estimate the relationship between diagnostic performance and parasitemia, should such data be made available. Finally, to address nonidentifiability issues, we assumed PCR to be the gold standard. However, varying limits of detection and the complexity of multiplex assays due to amplification issues, parasitemia, and laboratory settings among other factors may weaken this assumption. Although sensitivity analyses revealed that our species-level estimates were robust to this assumption, future work could jointly estimate the diagnostic performance of both methods, providing a more complete characterization of misdiagnosis of Plasmodium spp. in settings with co-circulation of multiple Plasmodium species.
This analysis estimates that the diagnostic performance of light microscopy for Plasmodium spp. may be limited in settings of cocirculation. Such misdiagnosis could have implications for malaria treatment and control, so more sensitive diagnostics, such as PCR, may be warranted in settings where Plasmodium spp. parasites routinely co-circulate.
Supplemental Materials
ACKNOWLEDGMENTS
We thank Parker Ladwig for helpful advice on conducting a systematic review.
REFERENCES
- 1.↑
World Health Organization , 2021. World Malaria Report 2021. Geneva, Switzerland: World Health Organization.
- 2.↑
Feachem RGA et al., 2019. Malaria eradication within a generation: ambitious, achievable, and necessary. Lancet 394: 1056–1112.
- 3.↑
Akala HM et al., 2021. Plasmodium interspecies interactions during a period of increasing prevalence of Plasmodium ovale in symptomatic individuals seeking treatment: an observational study. Lancet Microbe 2: e141–e150.
- 4.↑
Whittaker C , Slater H , Nash R , Bousema T , Drakeley C , Ghani AC , Okell LC , 2021. Global patterns of submicroscopic Plasmodium falciparum malaria infection: insights from a systematic review and meta-analysis of population surveys. Lancet Microbe 2: doi: S2666-5247(21)000550.
- 5.↑
Singh B , Sung LK , Matusop A , Radhakrishnan A , Shamsul SS , Cox-Singh J , Thomas A , Conway DJ , 2004. A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 363: 1017–1024.
- 6.↑
Barber BE , William T , Grigg MJ , Yeo TW , Anstey NM , 2013. Limitations of microscopy to differentiate Plasmodium species in a region co-endemic for Plasmodium falciparum, Plasmodium vivax and Plasmodium knowlesi. Malar J 12: 8.
- 7.↑
Anon , 2015. Guidelines for the Treatment of Malaria. 3rd ed. Geneva, Switzerland: World Health Organization.
- 8.↑
Kantele A , Jokiranta TS , 2011. Review of cases with the emerging fifth human malaria parasite, Plasmodium knowlesi. Clin Infect Dis 52: 1356–1362.
- 9.↑
Blumberg S , Lloyd-Smith JO , 2013. Comparing methods for estimating R0 from the size distribution of subcritical transmission chains. Epidemics 5: 131–145.
- 10.↑
Barber BE , William T , Dhararaj P , Anderios F , Grigg MJ , Yeo TW , Anstey NM , 2012. Epidemiology of Plasmodium knowlesi malaria in north-east Sabah, Malaysia: family clusters and wide age distribution. Malar J 11: 401.
- 11.↑
Baum E et al., 2016. Common asymptomatic and submicroscopic malaria infections in Western Thailand revealed in longitudinal molecular and serological studies: a challenge to malaria elimination. Malar J 15: 333.
- 12.↑
Chua KH , Lim SC , Ng CC , Lee PC , Lim YAL , Lau TP , Chai HC , 2015. Development of high resolution melting analysis for the diagnosis of human malaria. Sci Rep 5: 15671.
- 13.↑
Cooper DJ et al., 2020. Plasmodium knowlesi malaria in Sabah, Malaysia, 2015–2017: ongoing increase in incidence despite near-elimination of the human-only Plasmodium species. Clin Infect Dis 70: 361–367.
- 14.↑
Cox-Singh J , Davis TME , Lee K-S , Shamsul SSG , Matusop A , Ratnam S , Rahman HA , Conway DJ , Singh B , 2008. Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Clin Infect Dis 46: 165–171.
- 15.↑
Goh XT , Lim YAL , Vythilingam I , Chew CH , Lee PC , Ngui R , Tan TC , Yap NJ , Nissapatorn V , Chua KH , 2013. Increased detection of Plasmodium knowlesi in Sandakan division, Sabah as revealed by PlasmoNexTM . Malar J 12: 264.
- 16.↑
Han TZ , Han KT , Aye KH , Hlaing T , Thant KZ , Vythilingam I , 2017. Comparison of microscopy and PCR for the detection of human Plasmodium species and Plasmodium knowlesi in southern Myanmar. Asian Pac J Trop Biomed 7: 680–685.
- 17.↑
Lubis IND , Wijaya H , Lubis M , Lubis CP , Divis PCS , Beshir KB , Sutherland CJ , 2017. Contribution of Plasmodium knowlesi to multispecies human malaria infections in North Sumatera, Indonesia. J Infect Dis 215: 1148–1155.
- 18.↑
Putaporntip C , Hongsrimuang T , Seethamchai S , Kobasa T , Limkittikul K , Cui L , Jongwutiwes S , 2009. Differential prevalence of Plasmodium infections and cryptic Plasmodium knowlesi malaria in humans in Thailand. J Infect Dis 199: 1143–1150.
- 19.↑
Yusof R , Lau YL , Mahmud R , Fong MY , Jelip J , Ngian HU , Mustakim S , Hussin HM , Marzuki N , Mohd Ali M , 2014. High proportion of knowlesi malaria in recent malaria cases in Malaysia. Malar J 13: 168.
- 20.↑
Zhou X , Huang J-L , Njuabe MT , Li S-G , Chen J-H , Zhou X-N , 2014. A molecular survey of febrile cases in malaria-endemic areas along China–Myanmar border in Yunnan province, People’s Republic of China. Parasite 21: 27.
- 21.↑
Rahim MAFA , Munajat MB , Idris ZM , 2020. Malaria distribution and performance of malaria diagnostic methods in Malaysia (1980–2019): a systematic review. Malar J 19: 395.
- 22.↑
Gonçalves L , Subtil A , de Oliveira MR , do Rosário V , Lee P-W , Shaio M-F , 2012. Bayesian latent class models in malaria diagnosis. PLoS One 7: e40633.
- 23.↑
Carpenter B , Gelman A , Hoffman MD , Lee D , Goodrich B , Betancourt M , Brubaker M , Guo J , Li P , Riddell A , 2017. Stan: a probabilistic programming language. J Stat Soft 76.
- 24.↑
Mueller I , Galinski MR , Baird JK , Carlton JM , Kochar DK , Alonso PL , del Portillo HA , 2009. Key gaps in the knowledge of Plasmodium vivax, a neglected human malaria parasite. Lancet Infect Dis 9: 555–566.
- 25.↑
Baird JK , Battle KE , Howes RE , 2018. Primaquine ineligibility in anti-relapse therapy of Plasmodium vivax malaria: the problem of G6PD deficiency and cytochrome P-450 2D6 polymorphisms. Malar J 17: 42.